
|
Patrick AMAR
Équipe Bioinfo, LRI, Univ. Paris Sud et CNRS UMR 8623
| History |
HSIM has been developed since 2002. The first published reference can be found in Journal of Biological Physics and Chemistry, 2004, Vol 4, Number 2, pp:50-63.
| Some Examples |
| Download HSIM |
|
|
|
|
|
|
|
|
|
| The stochastic automaton HSIM |
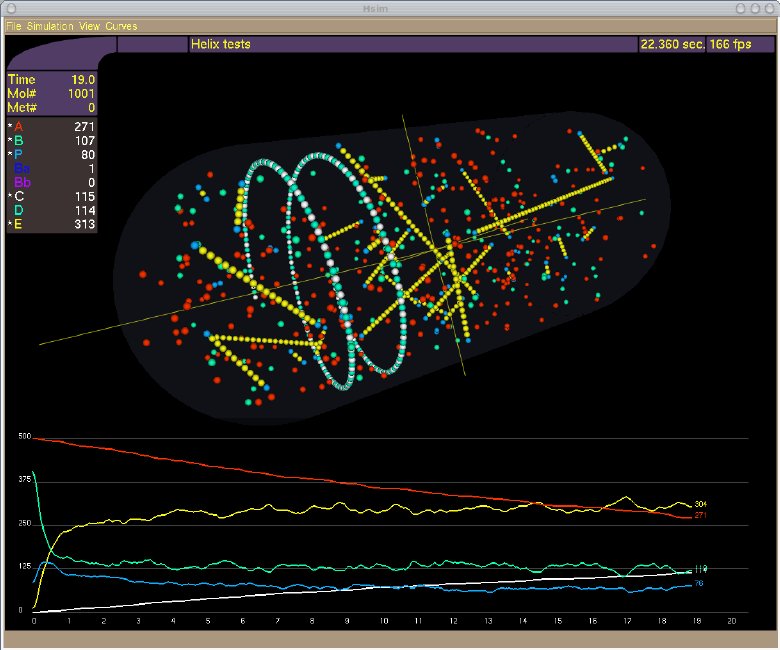
The simulator, HSIM, is a stochastic automaton driven by reaction rules between molecules.
In essence, each molecule is represented by a record that includes its type, its position, its size and a list of links to certain other molecules. HSIM keeps track of each molecule in real time from the computer point of view. The basic principle is that time is sliced into consecutive steps or generations, and in each generation the rules are applied to every molecule. These rules mimic the chemical reactions between molecules in a real system. The generation time is set to 100 microseconds, which corresponds to the average time for a protein to move a distance of 10 nanometers (of the order of its diameter) in vivo.
Metabolites diffuse faster than proteins, to take account of their smaller size, they are represented in HSIM by a sphere of reduced size with a greater diffusion speed. During a generation, the following processes are applied to all the molecules:
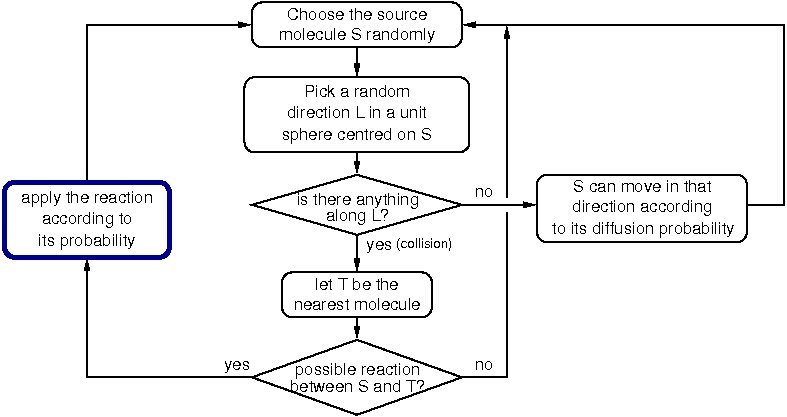
When all the molecules in the cell have been processed, the generation is completed and a new one begins. In HSIM the computer time is proportional to the total number of molecules and not to the size of the simulated space or the number of types of molecules.
One important point is that models in HSIM are additive: different models can be merged by simply merging their configuration files. If there are interactions between the models, HSIM will take them into account.
Each rule has an associated probability which corresponds to the kinetics of the reaction. For each kind of molecule, the maximum number of links to each other kind of molecule must be specified to allow the association rules to be functional.
title = "Enzymatic Reaction";
geometry = 120:40; // 1.2 x 0.4 nm
molecule s1, s2;
molecule E1;
size (s1) = 0.1;
size (s2) = 0.1;
speed (E1) = 0.1;
maxlinks (E1) = s1(1), s2(1);
maxlinks (s1) = E1(1);
maxlinks (s2) = E1(1);
E1 + s1 -> E1 * s1 [0.4]; // E1 captures its substrate
E1 * s1 -> E1 + s1 [1e-3]; // reverse reaction
E1 * s1 -> E1 * s2 [0.01]; // catalyse s1 -> s2
E1 * s2 -> E1 + s2 [0.01]; // E1 releases the product
init (30, E1);
init (1000, s1);
| Quick HSIM User Manual |
| -h | print this help. |
| -H | longer help (with interactive controls). |
| -b file | batch mode (no OpenGL display). |
| -bd file | batch mode (without diffusion phase). |
| -C file | count each reaction and write it in 'file'. |
| -m num | set the duration of the simulation (number of seconds of simulated time). |
| -q | quiet (no display at all). |
| -v | prints the rules on stderr. |
| -r num | initialise the random number generator. |
| -ra | randomly initialise the random number generator. |
| -R | display the rules. |
| -fs | display in full screen mode. |
| -s | 3D stereo mode. |
| -f file | use 'file' as configuration file. |
| -l file | load the simulation snapshot 'file' (infers the configuration). |
| -w | reload periodically the snapshot (watch file). |
| -g WxH | set the cell width and height. |
| -i num | set the number of generations between two histograms display. |
| -c MOL=num | add to the initial population of MOL 'num' more copies. |
| Keyboard controls | |
| a | show all the molecules (even those not linked) |
| b | show the backbone of the assemblies |
| d | toggle diffusion only / diffusion and reaction |
| D | set the length of the simulation in seconds |
| g | toggle concentration curves / assemblies histogram |
| h, ? | show this help |
| i | save the current display in a PNG image file |
| l | load a previously saved simulation |
| m | start / stop recording a movie of the simulation |
| n | normalise the scale for displaying the concentration curves |
| + | increase the scale factor |
| - | decrease the scale factor |
| q, Escape | exit the program |
| r | show / hide the links between bound molecules |
| R | show / hide the rules |
| s | toggle the 3D stereoscopic mode switch |
| S | save the current state of the simulator into a file |
| Tab | start/stop the simulation |
| Return | toggle display rate |
| Backspace | focus to the center of the cell |
| Mouse controls | |
| Left Drag | rotate around the X and Y axis |
| RIght Drag | change the aperture angle |
| Left Press | select a molecule to be the new center of rotation |
| Ctrl+Left Press | select an assembly to be shown |
| Mid Press | show a menu |
| title = "model name"; | name of the model |
| speed (mt) = prob; | diffusion speed expressed as a probability |
| size (mt) = num; | diameter of a molecule type in 10 mn unit. |
| geometry = lengh:diameter; | size of the cell in 10 mn units. |
| display (mt1, ..., mtN); | show the concentration curves of the species list. |
| asm name = (mt1, ..., mtN); | give the name name to all the assemblies containing the species list. |
| maxlinks (mt) = mt1 (nl1), ..., mtn (nln); | set the maximum number of links for species mt. |
| molecule descr1, ..., descrn | declare the species descr1, ..., descrn as cytosolic molecules where descri is 'mt [max link count] [hide] [inactive]' |
| membrane descr1, ..., descrn | declare the species descr1, ..., descrn as membrane molecules |
| species mt1, ..., mtn | declare the species as cytosolic molecules treated as an homogeneous population |
| init (#copies, mt); | fill the compartment with #copies copies of species mt. |
| init (conc uM, mt); | fill the compartment with conc micromolar of species mt. |
| init (conc mM, mt); | fill the compartment with conc millimolar of species mt. |
| surface (#copies, mt); | put #copies copies of the membrane species mt on one pole of the compartment membrane. |
| mt1 + mt2 -> mt3 + mt4 [prob]; | mt1 reacts with mt2 with probability prob; |
| mt1 + mt2 -> mt3 * mt4 [prob]; | mt1 reacts with mt2 with probability prob and forms a complex where mt1 become mt3 and mt2 become mt4. |
| mt1 * mt2 -> mt3 + mt4 [prob]; | the complex mt1 / mt2 dissociates with probability prob and mt1 become mt3 and mt2 become mt4. |
| mt1 * mt2 -> mt3 * mt4 [prob]; | the complex mt1 / mt2 reacts with probability prob to transform mt1 to mt3 and mt2 to mt4. |
Each molecule type of the left side of a rule can be more specific than simply the species name. The binding context can be expressed with this syntax:
| mt | an instance of molecule type mt, bound or not to any other molecule |
| {mt1}mt | an instance of molecule type mt which is already bound to a instance of molecule type mt1 |
| {~mt1}mt | an instance of molecule type mt which is not bound to a instance of molecule type mt1 |
geometry = 60:60;
molecule GOD; // Glucose oxydase. Km = 30 mM, Kcat = 337
species gluc, h2o2;
GOD (gluc -> h2o2) Km = 30 mM; Kcat = 337;
To implement this kind of reaction, HSIM use 3 standard rules and compute their
probabilities to match the Km and Kcat values:
GOD + gluc -> GODgl [0.04884]
GODgl -> GOD + gluc [0.8]
GODgl -> GOD + h2o2 [0.0674]